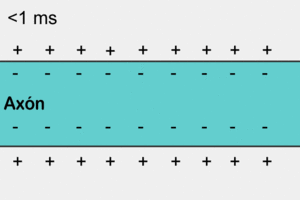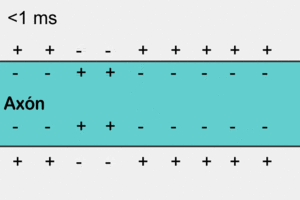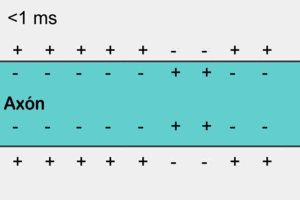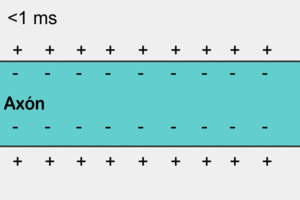
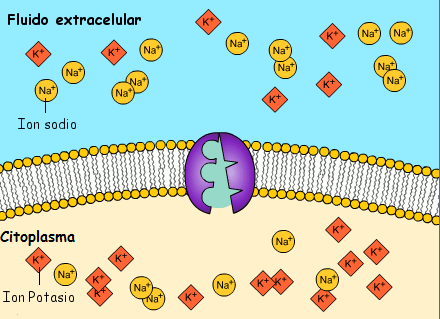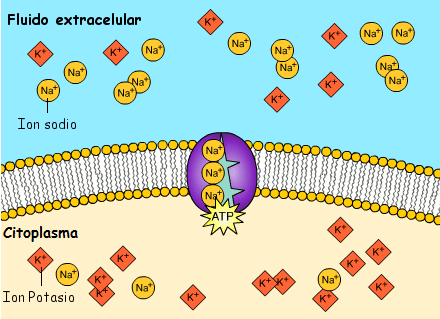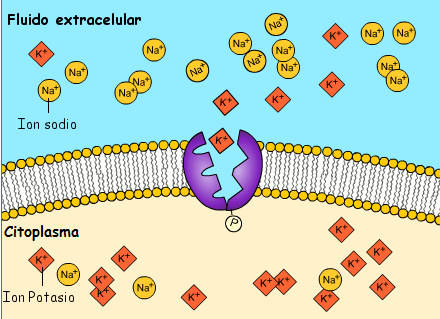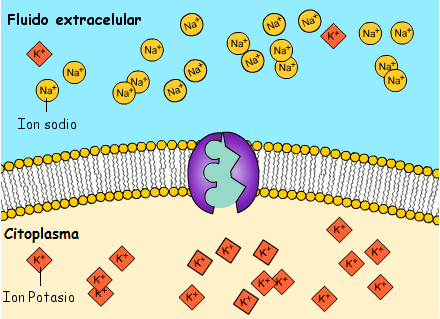
La concentración de potasio es grande dentro de la membrana de una fibra nerviosa, pero muy baja fuera de esta. A medida que lo hacen transportan cargas eléctricas positivas hacia el exterior, generando de esta manera electropositividad fuera de la membrana y electronegatividad en el interior debido a los aniones negativos que permanecen detrás y que no difunden hacia fuera con el potasio.
En la fibra nerviosa normal del mamífero la diferencia de potencial es de aproximadamente 94 mV, con negatividad en el interior de la membrana de la fibra.
El nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana se denomina potencial de Nernst para ese ion: la magnitud del potencial de Nernst viene determinada por el cociente de las concentraciones de ese ion específico en los dos lados de la membrana. Cuanto mayor es este cociente, mayor es la tendencia del ion a difundir en una dirección y, por tanto, mayor será el potencial de Nernst necesario para impedir la difusión neta adicional.
La ecuación de Goldman o ecuación de Goldman-Hodgkin-Katz, da el potencial de membrana calculado en el interior de la membrana cuando participan dos iones positivos univalentes, sodio (Na+) y potasio (K+), y un ion negativo univalente, cloruro (Cl–).
POTENCIAL DE MEMBRANA
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV.
Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.
La bomba sodio-potasio es una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular. El denominado canal de potasio de dominios de poro en tándem, o canal de «fuga» de potasio (K+), en la membrana nerviosa a través de la que pueden escapar iones potasio incluso en una célula en reposo.
El potencial de Nernst que corresponde a este cociente es de –94 mV porque el logaritmo de 35 es 1,54, y 1,54 multiplicado por –61 mV es –94 mV. Por tanto, si los iones potasio fueran el único factor que genera el potencial en reposo, el potencial en reposo en el interior de la fibra sería igual a –94 mV, como se muestra en la figura.
Potencial de acción de las neuronas
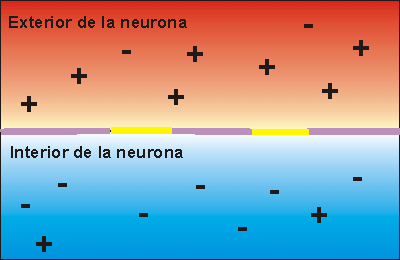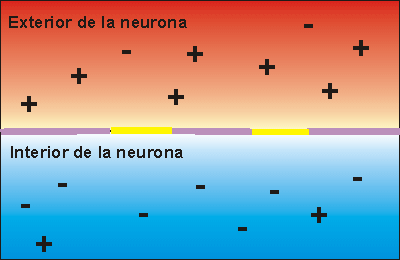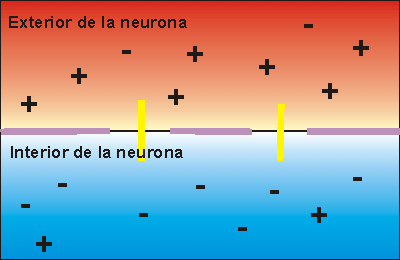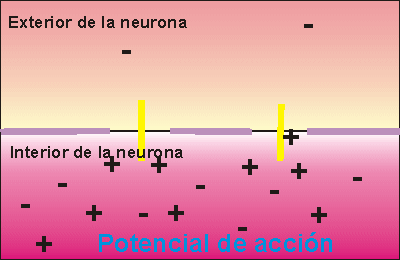
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo.
Activación e inactivación del canal de sodio activado por el voltaje
El canal de sodio activado por el voltaje en tres estados distintos. Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. La parte superior izquierda de la figura representa el estado de estas dos compuertas en la membrana en reposo normal, cuando el potencial de membrana es de –90 mV. En este estado la compuerta de activación está cerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra a través de estos canales de sodio.
Activación del canal de sodio Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje (habitualmente algún punto entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta. Durante este estado activado, los iones sodio pueden atravesar el canal, aumentando la permeabilidad de la membrana al sodio hasta 500 a 5.000 veces.
Durante el estado de reposo, antes de que comience el potencial de acción, la conductancia a los iones potasio es 50 a 100 veces mayor que la conductancia a los iones sodio. Esta disparidad se debe a una fuga mucho mayor de iones potasio que sodio a través de los canales de fuga. Sin embargo, al inicio del potencial de acción se activan instantáneamente los canales de sodio y dan lugar a un aumento de la conductancia al sodio de 5.000 veces. Después el proceso de inactivación cierra los canales de sodio en otra fracción de milisegundo.
Un ciclo de retroalimentación positiva abre los canales de sodio Primero, siempre que no haya alteraciones de la membrana de la fibra nerviosa, no se produce ningún potencial de acción en el nervio normal. Sin embargo, si algún episodio produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje. Esta situación permite la entrada rápida de iones sodio, lo que produce una elevación adicional del potencial de membrana y abre aún más canales de sodio activados por el voltaje y permite que se produzca una mayor entrada de iones sodio hacia el interior de la fibra.
Umbral para el inicio del potencial de acción No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva que se ha descrito en el párrafo anterior. Esto se produce cuando el número de iones sodio que entran en la fibra supera al número de iones potasio que salen. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV.
Ritmicidad de algunos tejidos excitables: descarga repetitiva Las descargas repetitivas autoinducidas aparecen normalmente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central. Estas descargas rítmicas producen: 1) el latido rítmico del corazón; 2) el peristaltismo rítmico de los intestinos, y 3) fenómenos neuronales, como el control rítmico de la respiración.
Las fibras grandes son mielinizadas y las pequeñas no mielinizadas. Un tronco nervioso medio contiene aproximadamente el doble de fibras no mielinizadas que mielinizadas.
Las células de Schwann depositan la vaina de mielina alrededor del axón de la siguiente manera: en primer lugar, la membrana de una célula de Schwann rodea el axón. Después, la célula de Schwann rota muchas veces alrededor del axón, depositando múltiples capas de membrana de la célula de Schwann que contiene la sustancia lipídica esfingomielina. Esta sustancia es un excelente aislante eléctrico que disminuye el flujo iónico a través de la membrana aproximadamente 5.000 veces. En la unión entre dos células de Schwann sucesivas a lo largo del axón permanece una pequeña zona no aislada de solo 2 a 3 μm de longitud en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón. Esta zona se denomina nódulo de Ranvier.
No se puede producir un nuevo potencial de acción en una fibra excitable mientras la membrana siga despolarizada por el potencial de acción precedente. El período durante el cual no se puede generar un segundo potencial de acción, incluso con un estímulo intenso, se denomina período refractario absoluto. Para las fibras nerviosas mielinizadas grandes este período es de aproximadamente 1/2.500 s.
