Las señales de entrada llegan a las neuronas través de las sinapsis situadas fundamentalmente en las dendritas neuronales, pero también en el soma celular. Según los diversos tipos de neuronas, las conexiones sinápticas procedentes de las fibras aferentes pueden ser tan solo unos cientos o llegar hasta 200.000.
La porción somática del sistema sensitivo, que transmite información sensitiva desde los receptores repartidos por la superficie de todo el cuerpo y desde algunas estructuras profundas.
Sistema nervioso autónomo, opera de forma paralela a su acción, estando encargado de controlar la musculatura lisa, las glándulas y otros sistemas corporales internos.
Una de las funciones más importantes del sistema nervioso consiste en elaborar la información que le llega de tal modo que dé lugar a las respuestas motoras y mentales adecuadas. El encéfalo descarta más del 99% de toda la información sensitiva que recibe por carecer de interés o de importancia. señales facilitadoras e inhibidoras procedentes de otras regiones del sistema nervioso tienen la capacidad de controlar la transmisión sináptica, a veces abriendo las sinapsis para efectuar la comunicación y en otras ocasiones cerrándolas. Además, algunas neuronas postsinápticas responden con un amplio número de impulsos de salida y otras lo hacen solo con unos pocos. La mayor parte del almacenamiento tiene lugar en la corteza cerebral, pero hasta las regiones basales del encéfalo y la médula espinal pueden conservar pequeñas cantidades de información. La acumulación de la información es el proceso que llamamos memoria, y también constituye una función de las sinapsis.
los procedimientos de deliberación del encéfalo comparan las experiencias sensitivas nuevas con los recuerdos acumulados; a continuación, estos últimos sirven para seleccionar la información sensitiva nueva que resulte más importante y encauzarla hacia las regiones correspondientes para el almacenamiento de la memoria a fin de permitir su uso en el futuro o hacia las regiones motoras para dar lugar a las respuestas corporales inmediatas.
La información recorre el sistema nervioso central sobre todo bajo la forma de potenciales de acción nerviosos, llamados simplemente impulsos nerviosos, a través de una sucesión de neuronas, una después de la otra.
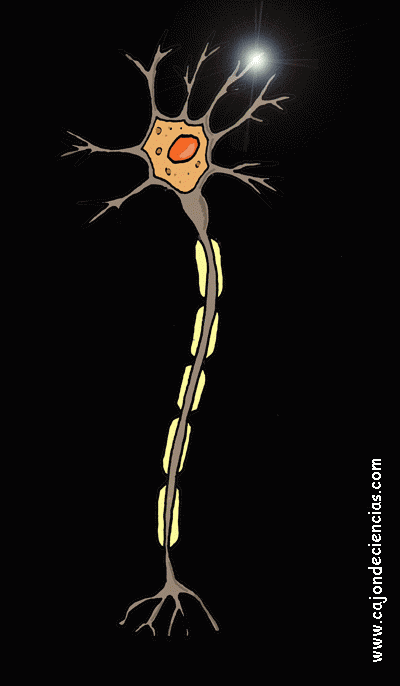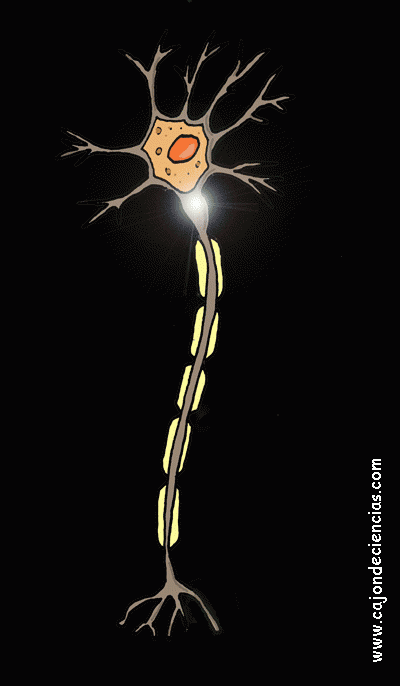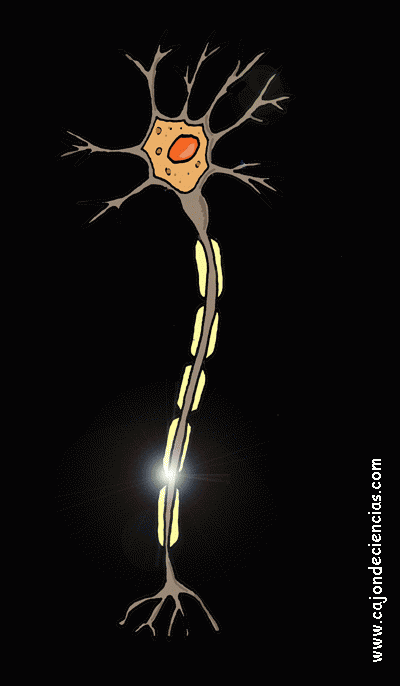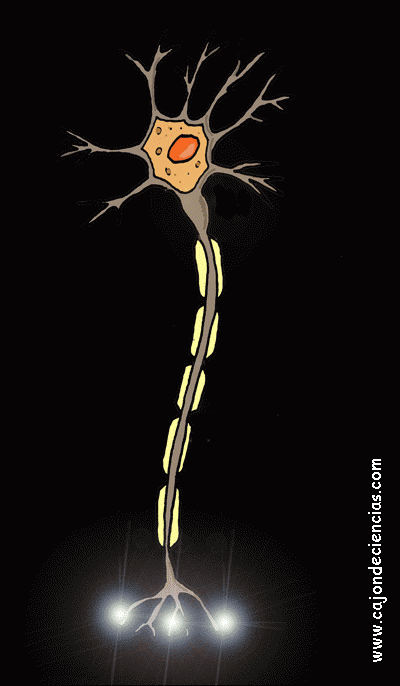
La mayoría de las sinapsis utilizadas para la transmisión de señales en el sistema nervioso central del ser humano son sinapsis químicas. En estas sinapsis, la primera neurona segrega un producto químico denominado neurotransmisor (a menudo llamado sustancia transmisora) a nivel de la terminación nerviosa, que a su vez actúa sobre las proteínas receptoras presentes en la membrana de la neurona siguiente para excitarla, inhibirla o modificar su sensibilidad de algún otro modo. Hasta hoy se han descubierto más de 40 neurotransmisores importantes.
En las sinapsis eléctricas los citoplasmas de las células adyacentes están conectados directamente por grupos de canales de iones llamados uniones en hendidura que permiten el movimiento libre de los iones desde el interior de una célula hasta el interior de la siguiente.
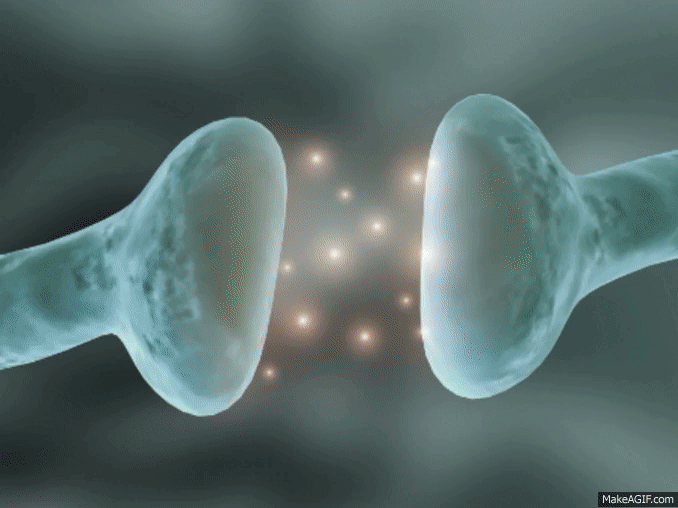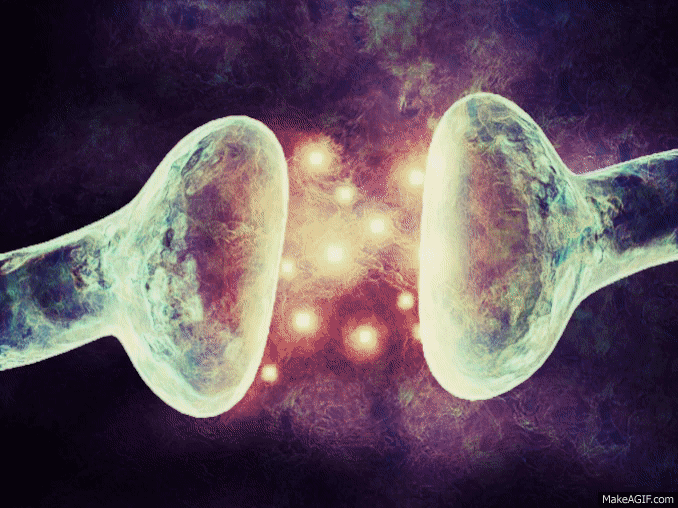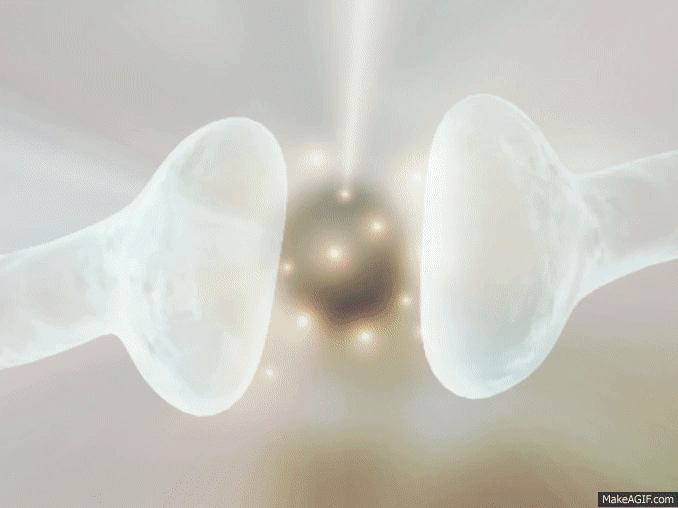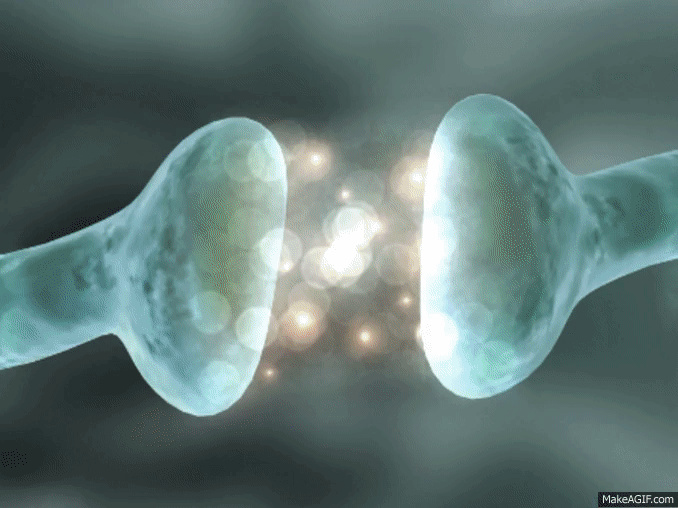
La membrana del terminal presináptico se llama membrana presináptica. Contiene una gran abundancia de canales de calcio dependientes de voltaje.
Los canales iónicos de la membrana neuronal postsináptica suelen ser de dos tipos: 1) canales catiónicos, cuya clase más frecuente deja pasar iones sodio cuando se abren, pero a veces también cumplen esta función con el potasio o el calcio, y 2) canales aniónicos, que permiten sobre todo el paso de los iones cloruro, pero también de minúsculas cantidades de otros aniones.
Existen diversos tipos de sistemas de segundo mensajero. Uno de los más frecuentes recurre a un grupo de proteínas llamadas proteínas G. El complejo de proteínas G inactivas está libre en el citosol y consta de difosfato de guanosina (GDP) más tres elementos: un componente alfa (α), que es la porción activadora de la proteína G, y unos componentes beta (β) y gamma (γ) que están pegados al componente α. Mientras el complejo de proteínas G está unido a GDP, permanece inactivo.
Las vesículas que se almacenan y liberan transmisores de molécula pequeña se reciclan continuamente y se utilizan una y otra vez. Una vez que se fusionan con la membrana sináptica y se abren para verter la sustancia transmisora, la membrana de la vesícula simplemente forma parte al principio de la membrana sináptica.
Los neuropéptidos se sintetizan de otro modo y tienen acciones que normalmente son lentas y en otros aspectos bastante diferentes de las que ejercen los transmisores de molécula pequeña. Las moléculas proteicas penetran a continuación en los espacios existentes en el retículo endoplásmico del soma y posteriormente en el aparato de Golgi, donde suceden dos cambios. se transportan por el axón en todas las direcciones hacia el extremo de las fibras nerviosas a través de la corriente axónica del citoplasma, viajando a una velocidad de tan solo unos pocos centímetros al día. Finalmente, estas vesículas vierten su contenido en los terminales neuronales como respuesta a los potenciales de acción de la misma manera que los transmisores de molécula pequeña. Sin embargo, la vesícula sufre una autólisis y no se reutiliza.
un potencial de membrana en reposo de unos –65 mV. Este potencial de membrana en reposo es un poco menos negativo que los – 90 mV existentes en las grandes fibras nerviosas periféricas y en las del músculo esquelético.
La entrada de cloruro más la salida de potasio elevan el grado de negatividad intracelular, lo que se denomina hiperpolarización. Un aumento de la negatividad por encima del potencial de membrana en reposo normal se denomina potencial postsináptico inhibidor (PPSI). efecto aditivo de los potenciales postsinápticos simultáneos mediante la activación de múltiples terminales situados en regiones muy espaciadas de la membrana neuronal se denomina sumación espacial.
