Por medio de los nervios ópticos las señales nerviosas abandonan la retina. Los tractos ópticos o cintillas ópticas se forman cuando en el quiasma óptico, las fibras procedentes de la mitad nasal de la retina cruzan hacia el lado opuesto, donde se unen a las fibras originadas en la retina temporal contraria. Después, se produce una sinapsis de las fibras de cada tracto óptico en el núcleo geniculado del tálamo, a partir del cual las fibras geniculocalcarinas se dirigen a través de la radiación óptica constituyendo el tracto geniculocalcarino, que se dirige hacia la corteza visual primaria en el área correspondiente a la cisura calcarina del lóbulo occipital medial.
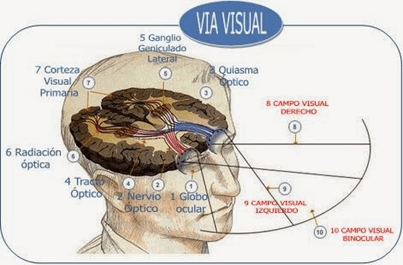
El núcleo geniculado lateral dorsal o cuerpo geniculado lateral está situado en el extremo dorsal del tálamo. El cual está compuesto por seis capas nucleares, las capas II, III y V (de ventral a dorsal) reciben señales desde la mitad lateral de la retina del mismo lado, mientras que las capas I, IV y VI las reciben de la mitad medial de la retina del ojo contralateral. Las capas I y II se llaman capas magnocelulares porque contienen neuronas grandes proporcionando una vía de conducción rápida hacia la corteza visual. En cambio, las capas III a VI se denominan capas parvocelulares porque poseen una gran cantidad de neuronas de tamaño pequeño a mediano. Esta estructura tiene las siguientes funciones:
- Transfiere la información visual desde el tracto óptico hacia la corteza visual a través de la radiación óptica o tracto geniculocalcarino
- Filtra la transmisión de los impulsos hacia la corteza visual
La corteza visual primaria o área visual I, se halla en el área de la cisura calcarina, extendiéndose desde el polo occipital hacia delante por la cara medial de cada corteza occipital. Es la estación terminal de las señales visuales directas procedentes de los ojos. La corteza visual primaria posee seis capas diferentes, la capa IV está entrelazada con las bandas de las columnas neuronales, cuya anchura mide unos 0,5 mm. La corteza visual posee una organización estructural formada por varios millones de columnas verticales de células neuronales, con un diámetro de 30 a 50 μm cada una.
La mácula es una zona a la que envía sus señales la fóvea de la retina. Las áreas visuales secundarias o áreas visuales de asociación, ocupan zonas laterales, anteriores, superiores e inferiores a la corteza visual primaria.
Después de abandonar la corteza visual primaria, la información referida a la visión se analiza por dos vías principales de las áreas visuales secundarias:
1ra vía. Análisis de la posición tridimensional, la forma global y el movimiento de los objetos: examina la posición tridimensional que ocupan los objetos visuales en el espacio que rodea al organismo y explora la forma física global de la escena visual. Los impulsos viajan en general hacia el área temporal media posterior y ascienden hacia la extensa corteza occipitoparietal
2da vía. Análisis de los detalles visuales y del color: se encarga de la identificación de las letras, la lectura, la determinación de la textura de los objetos, de sus colores detallados, y de descifrar lo que es y lo que significa un objeto a partir de toda esta información.
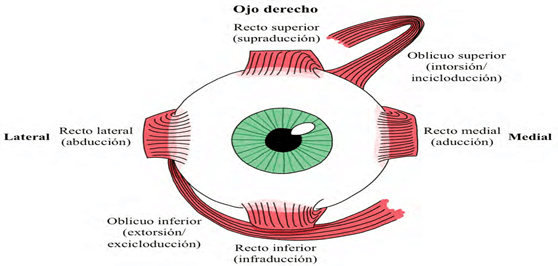
Los movimientos oculares están controlados por tres pares de músculos: Los rectos medial y lateral, los rectos superior e inferior y los oblicuos superior e inferior.
Los mecanismos neuronales son encargados de la fijación de los ojos, permitiendo el mecanismo voluntario de fijación (controlado por controlados por un campo cortical) y el mecanismo involuntario de fijación.
Para que las percepciones visuales resulten más elocuentes, generalmente se fusionan entre sí las imágenes visuales de ambos ojos según los puntos correspondientes de las dos retinas.

El mecanismo neuronal para la estereopsia o percepción de profundidad se basa en el hecho de que algunas de las vías integradas por las fibras que van desde la retina hacia la corteza visual se apartan cada lado del trayecto central, ciertas vías ópticas procedentes de ambos ojos coinciden exactamente para los objetos a 2 m de distancia, mientras que otro grupo diferente lo hace para los que están situados a 25 m.
El ojo está inervado por fibras nerviosas simpáticas y parasimpáticas. Las fibras preganglionares parasimpáticas nacen en el núcleo de Edinger-Westphal y a continuación viajan en el tercer par hasta el ganglio ciliar, que se halla justo detrás del ojo. Los axones preganglionares hacen sinapsis con las neuronas parasimpáticas posganglionares, las cuales envían sus fibras hacia el globo ocular a través de los nervios ciliares, y estos excitan el músculo ciliar (controla el enfoque del cristalino) y el esfínter del iris (contrae la pupila).
Para tener un buen nivel de agudeza visual es fundamental el mecanismo de acomodación del sistema ocular de lentes. La acomodación deriva de la contracción o relajación del músculo ciliar del ojo. La acomodación del cristalino está regulada por un mecanismo de retroalimentación negativo que corrige automáticamente su poder dióptrico para lograr el mayor grado de agudeza visual.
Miosis es disminución de la apertura pupilar, provocada por la excitación del músculo esfínter de la pupila que es estimulado por nervios parasimpáticos. Midriasis es lo contrario, ocurre con la estimulación de los nervios simpáticos que excita las fibras radiales del iris.

El reflejo pupilar fotomotor es cuando las pupilas se contraen a causa de la luz, la cual incide sobre la retina, parte de las señales activadas se dirigen desde los nervios ópticos hasta los núcleos pretectales. La función del reflejo fotomotor consiste en ayudar al ojo a la adaptación rápida a unas condiciones lumínicas cambiantes.
