El principal objetivo de la microcirculación es el transporte de nutrientes hacia los tejidos y la eliminación de los restos celulares. Cada tejido controla su propio flujo sanguíneo dependiendo de sus necesidades individuales. La circulación periférica de todo el organismo tiene alrededor de 10.000 millones de capilares con una superficie total estimada de 500-700 m2.
Cada arteria nutricia que entra en un órgano se ramifica seis u ocho veces antes de que las arterias sean suficientemente pequeñas para denominarse arteriolas, tienen diámetros internos de solo 10-15 μm. Las arteriolas se ramifican entre dos y cinco veces, alcanzando diámetros de 5 a 9 μm. Las metaarteriolas no tienen una capa muscular continua, sino fibras musculares lisas rodeando el vaso en puntos intermitentes.
El esfínter precapilar es el punto en el que cada capilar verdadero se origina de una metaarteriola. Las vénulas son mayores que las arteriolas y tienen una capa muscular mucho más débil. A pesar de ello, la presión de las vénulas es mucho menor que la de las arteriolas, por lo que las vénulas aún pueden contraerse considerablemente, a pesar de su capa muscular débil.

La pared capilar está compuesta por una capa unicelular de células endoteliales y rodeada por una membrana basal muy fina en el exterior del capilar. El grosor total de la pared capilar es de solo unas 0,5 μm, el diámetro interno del capilar es de 4-9 μm. Existe un espacio intercelular, que es un canal curvo a modo de hendidura fina-poro que descansa en la parte superior de la figura entre células endoteliales adyacentes, cada espacio tiene pliegues cortos de inserciones de proteínas que mantienen unidas las células endoteliales, pero entre esos pliegues puede filtrarse libremente el líquido a través del espacio, tiene una anchura de 6-7 nm. Se encuentran situados solo en los bordes de las células endoteliales, no representan más de 1/1.000 de la superficie total de la pared capilar.
En las células endoteliales también hay muchas vesículas de plasmalema o cavéolas (pequeñas cuevas), las cuales se forman a partir de oligómeros de proteínas llamadas caveolinas que están asociadas con moléculas de colesterol y esfingolípidos. Tiene una función en la endocitosis y en la transcitosis de macromoléculas en el interior de las células endoteliales
1. En el cerebro, las uniones entre las células endoteliales capilares son principalmente uniones estrechas que permiten la entrada y salida de moléculas muy pequeñas como agua, oxígeno y dióxido de carbono en los tejidos cerebrales.
2. En el hígado sucede lo contrario. Los espacios entre las células endoteliales capilares son aperturas amplias, por lo que casi todas las sustancias disueltas en el plasma, incluidas las proteínas plasmáticas, pueden pasar de la sangre a los tejidos hepáticos.
3. Los poros de las membranas capilares gastrointestinales tienen un tamaño intermedio entre las de los músculos y las del hígado.
4. En los capilares glomerulares del riñón se abren numerosas membranas ovales, denominadas fenestraciones, que atraviesan en todo su trayecto las células endoteliales, por lo que pueden filtrarse cantidades enormes de moléculas pequeñas e iones (pero no las moléculas grandes de las proteínas plasmáticas) a través de los glomérulos sin tener que pasar a través de los espacios situados entre las células endoteliales.
La sangre no fluye continuamente a través de los capilares, sino que lo hace de forma intermitente apareciendo y desapareciendo cada pocos segundos o minutos. La causa de esta intermitencia es el fenómeno conocido como vasomotilidad, lo que significa la contracción intermitente de las metaarteriolas y esfínteres precapilares.

El factor más importante que afecta al grado de apertura y cierre de las metaarteriolas y de los esfínteres precapilares, y que se ha descubierto hasta la fecha, es la concentración de oxígeno en los tejidos. Cuando la velocidad de utilización del oxígeno por el tejido es mayor, de forma que la concentración de oxígeno tisular disminuye por debajo de lo normal, se activan los períodos intermitentes del flujo sanguíneo capilar más a menudo y la duración de cada período del flujo es más prolongada, con lo que se permite que la sangre capilar transporte mayores cantidades de oxígeno hacia los tejidos. Hay una velocidad media del flujo sanguíneo a través de cada lecho capilar tisular, una presión capilar media dentro de los capilares y una velocidad de transferencia media de las sustancias entre la sangre de los capilares y el líquido intersticial circundante.
El medio más importante por el cual se transfieren las sustancias entre el plasma y el líquido intersticial es la difusión. Gran cantidad de moléculas de agua y partículas disueltas que entran y salen a través de la pared capilar, permitiendo la mezcla continua entre el líquido intersticial y el plasma. La difusión es consecuencia del movimiento térmico de las moléculas de agua y de otras sustancias disueltas en el líquido, con las distintas moléculas e iones desplazándose primero en una dirección y luego en otra, rebotando aleatoriamente en cada una de ellas. Si una sustancia es liposoluble (oxígeno y el dióxido de carbono), difunde directamente a través de las membranas celulares del capilar sin tener que atravesar los poros.
Muchas sustancias que necesitan los tejidos son solubles en agua pero no pueden pasar a través de las membranas lipídicas de las células endoteliales; estas sustancias son las propias moléculas de agua, los iones sodio y cloruro y la glucosa. la velocidad con la que difunden las moléculas de agua a través de la membrana capilar es unas 80 veces mayor que la velocidad con la que el propio plasma fluye linealmente por el capilar. La profundidad de los espacios intercelulares capilares, 6 a 7 nm, es unas 20 veces el diámetro de la molécula de agua, que es la molécula más pequeña que normalmente atraviesa los poros de los capilares. , las membranas de los sinusoides del capilar hepático son tan permeables que incluso las proteínas plasmáticas atraviesan esas paredes casi tan fácilmente como el agua y otras sustancias. Además, la permeabilidad de la membrana glomerular renal para el agua y los electrólitos es unas 500 veces mayor que la permeabilidad de los capilares musculares, aunque no para las proteínas plasmáticas; para estas proteínas, la permeabilidad de los capilares es muy pequeña, como en otros órganos y tejidos.
La velocidad “neta” de difusión de una sustancia a través de cualquier membrana es proporcional a la diferencia de concentración de la sustancia entre los dos lados de la membrana. Las velocidades de difusión a través de la membrana capilar de las sustancias más importantes para la nutrición son tan grandes que solo diferencias pequeñas de concentración son suficientes para que el transporte entre el plasma y el líquido intersticial sea más que adecuado.
Una sexta parte del volumen total del organismo consiste en espacios entre las células, que colectivamente se conoce como el intersticio. El líquido de estos espacios se denomina líquido intersticial.
La estructura del intersticio contiene dos tipos principales de estructuras sólidas:
1) haces de fibras de colágeno
2) filamentos de proteoglucano. Los haces de las fibras de colágeno recorren largas distancias en el intersticio. Son muy fuertes, por lo que proporcionan la mayor parte de la fuerza tensional de los tejidos.
El líquido del intersticio deriva por filtración y difusión de los capilares. Contiene casi los mismos componentes que el plasma, excepto por concentraciones mucho más bajas de proteínas, porque las proteínas no atraviesan los poros de los capilares. El líquido intersticial queda atrapado principalmente en los diminutos espacios que hay entre los filamentos de proteoglucanos.
Aunque casi todo el líquido del intersticio está atrapado dentro del gel tisular, a veces también hay pequeños riachuelos de líquido «libre» y pequeñas vesículas de líquido libre, lo que significa que carece de moléculas de proteoglucano y, por tanto, puede fluir libremente. Cuando se inyecta un colorante en la sangre circulante, a menudo puede verse fluir a través del intersticio en los riachuelos pequeños que circulan habitualmente por las superficies de las fibras de colágeno o en las superficies de las células. La cantidad de líquido «libre» presente en los tejidos normales es pequeña, mucho menor del 1%. Por el contrario, cuando se desarrolla edema en los tejidos, estos bolsillos y riachuelos pequeños de líquido libre se expanden mucho hasta que la mitad o más del líquido del edema comienza a fluir libremente, independientemente de los filamentos de proteoglucano.
La presión hidrostática en los capilares tiende a empujar el líquido y a las sustancias disueltas a través de los poros capilares dentro de los espacios intersticiales. Por el contrario, la presión osmótica provocada por las proteínas plasmáticas (lo que se conoce como presión coloidosmótica) tiende a provocar el movimiento del líquido por ósmosis desde los espacios intersticiales hacia la sangre. Esta presión osmótica ejercida por las proteínas plasmáticas normalmente previene la pérdida significativa de volumen de líquido desde la sangre hacia los espacios intersticiales. El sistema linfático también tiene su importancia, al devolver a la circulación las pequeñas cantidades del exceso de proteína y líquido que se pierde desde la sangre hacia los espacios intersticiales. En el resto de este capítulo comentaremos los mecanismos que controlan a la vez la filtración capilar y el flujo linfático para regular los volúmenes respectivos de plasma y líquido intersticial.
Cuatro fuerzas principales que determinan si el líquido saldrá de la sangre hacia el líquido intersticial o en dirección contraria. Estas fuerzas, denominadas “fuerzas de Starling”, son:
1. La presión capilar (Pc), que tiende a forzar la salida del líquido a través de la membrana capilar.
2. La presión del líquido intersticial (Pif), que tiende a forzar la entrada del líquido a través de la membrana capilar cuando la Pif es positiva, pero fuerza la salida cuando la Pif es negativa.
3. La presión coloidosmótica del plasma en el capilar (Πp), que tiende a provocar ósmosis de líquido hacia el interior a través de la membrana capilar.
4. La presión coloidosmótica del líquido intersticial (Πif), que tiende a provocar la ósmosis del líquido hacia el exterior a través de la membrana capilar.
Ernest Starling señaló hace más de un siglo que, en condiciones normales, existe un estado cercano al equilibrio en la mayoría de los capilares, es decir, que la cantidad de líquido que se filtra de los extremos arteriales de los capilares hacia el exterior es casi exactamente igual a la de líquido que vuelve a la circulación mediante absorción. El ligero desequilibrio que se produce explica el líquido que puede volver a la circulación a través de los vasos linfáticos. En el diagrama siguiente se muestran los principios de equilibrio de Starling. Para su elaboración se obtiene la media de las presiones de los capilares arteriales y venosos para calcular la media de la presión capilar funcional a lo largo de todo el capilar. Se calcula que esta presión capilar funcional media es de 17,3 mmHg. Si la presión capilar media aumenta por encima de 17 mmHg aumenta también la fuerza neta que tiende a causar la filtración de líquido en los espacios tisulares. Es decir, un aumento de 20 mmHg de la presión capilar media provoca un incremento de la presión de filtración de 0,3 mmHg a 20,3 mmHg, con lo que se consigue una filtración neta hasta 68 veces mayor que la que ocurre normalmente en los espacios intersticiales. Para prevenir la acumulación del exceso de líquido en estos espacios se requiere 68 veces más flujo de líquido normal en el sistema linfático, una cantidad que es 2 a 5 veces mayor que la que pueden eliminar los vasos linfáticos. En consecuencia, comenzará a acumularse el líquido en los espacios intersticiales y se producirá edema.
El sistema linfático representa una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. Es más, los vasos linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de estas podrá ser eliminada por absorción directamente hacia los capilares sanguíneos. Este retorno de las proteínas a la sangre desde los espacios intersticiales es una función esencial sin la cual moriríamos en 24 h. Casi todos los tejidos del organismo tienen vasos linfáticos especiales que drenan el exceso de líquido directamente desde los espacios intersticiales.
Hay algunas excepciones, como las porciones superficiales de la piel, el sistema nervioso central y el endomisio de músculos y huesos. Casi todos los tejidos del organismo tienen vasos linfáticos especiales que drenan el exceso de líquido directamente desde los espacios intersticiales.
La linfa de la mitad izquierda de la cabeza, el brazo izquierdo y algunos territorios del tórax entra en el conducto torácico antes de que se vacíe en las venas.

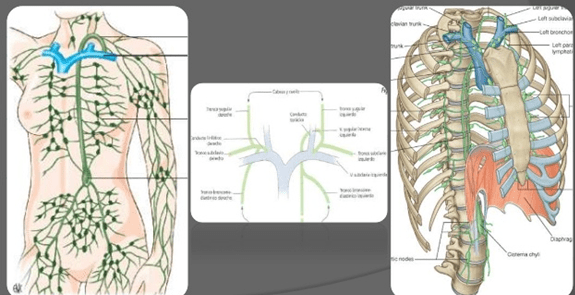
La linfa del lado derecho del cuello y la cabeza, el brazo derecho y algunos territorios del tórax derecho entra en el conducto torácico derecho (mucho menor que el conducto torácico izquierdo), que se vacía en el sistema venoso en la unión de la vena subclavia derecha y la vena yugular interna.

La linfa deriva del líquido intersticial que fluye en los linfáticos, por lo que la linfa que entra primero en los vasos linfáticos terminales tiene casi la misma composición que el líquido intersticial. La concentración de proteínas en el líquido intersticial de la mayoría de los tejidos alcanza un promedio de 2 g/dl y la concentración de proteínas del flujo linfático que procede de estos tejidos es aproximada a este valor. La linfa formada en el hígado tiene una concentración de proteínas hasta de 6 g/dl y la linfa formada en el intestino tiene una concentración de proteínas hasta de 3-4 g/dl. Como aproximadamente dos tercios de toda la linfa procede normalmente del hígado y los intestinos, la linfa del conducto torácico, que es una mezcla de linfa de todas las áreas del organismo, tiene una concentración de proteínas en torno a 3-5 g/dl.
El capilar linfático terminal también puede bombear la linfa, efecto que se suma al bombeo producido en los vasos linfáticos mayores. Como hemos visto antes en este mismo capítulo, las paredes de los linfáticos están íntimamente adheridas a las células tisulares circundantes, mediante sus filamentos de anclaje. Por tanto, cada vez que entra un exceso de líquido en el tejido y provoca la hinchazón tisular, los filamentos de anclaje tiran de la pared de los capilares linfáticos y el flujo entrará en el capilar linfático terminal a través de los espacios situados entre las células endoteliales. Después, la presión del interior del capilar aumenta cuando se comprime el tejido y se provoca la superposición de los bordes de las células endoteliales, que se cierran a modo de válvulas. Por tanto, la presión empuja la linfa a través de los espacios intercelulares hacia el linfático colector, y no hacia atrás. Las células endoteliales de los capilares linfáticos también contienen algunos filamentos contráctiles de actomiosina.
