Un principio fundamental de la función circulatoria es que la mayoría de los tejidos tienen la capacidad de controlar su propio flujo sanguíneo local en proporción a sus necesidades metabólicas concretas. Algunas de las necesidades específicas de flujo sanguíneo en los tejidos incluyen aspectos como:
1. Aporte de oxígeno a los tejidos.
2. Aporte de otros nutrientes, como glucosa, aminoácidos y ácidos grasos.
3. Eliminación de dióxido de carbono de los tejidos.
4. Eliminación de iones hidrógeno de los tejidos.
5. Mantenimiento de las concentraciones adecuadas de iones en los tejidos.
6. Transporte de varias hormonas y otras sustancias a los distintos tejidos.

El control del flujo sanguíneo local se puede dividir en dos fases:
1) control a corto plazo. Consigue con cambios rápidos de la vasodilatación o vasoconstricción local de las arteriolas, metaarteriolas y esfínteres precapilares, que se producen en segundos o minutos para proporcionar con gran rapidez el mantenimiento del flujo sanguíneo tisular local apropiado.
2) control a largo plazo. Significa cambios controlados lentos del flujo en un período de días, semanas o incluso meses. En general, estos cambios a largo plazo proporcionan un control aún mejor del flujo en proporción a las necesidades de los tejidos. Estos cambios se producen como consecuencia del incremento o descenso del tamaño físico y del número de vasos sanguíneos que nutren los tejidos.
Según la teoría vasodilatadora, cuanto mayor sea el metabolismo o menor sea la disponibilidad de oxígeno o de algunos otros nutrientes en un tejido, mayor será la velocidad de formación de sustancias vasodilatadoras en las células de ese tejido. Se cree que estas sustancias vasodilatadoras difunden a través de los tejidos hacia los esfínteres precapilares, las metaarteriolas y las arteriolas para provocar la dilatación. Se han propuesto varias sustancias vasodilatadoras diferentes, como adenosina, dióxido de carbono, compuestos con fosfato de adenosina, histamina, iones potasio e iones hidrógeno.
Las sustancias pueden liberarse del tejido en respuesta a la deficiencia de oxígeno. Por ejemplo, en los experimentos se ha demostrado que la reducción de la disponibilidad de oxígeno provoca la liberación tanto de adenosina como de ácido láctico (que contiene iones hidrógeno) en los espacios libres entre las células tisulares; estas sustancias provocan una vasodilatación aguda a corto plazo y, por tanto, son responsables, o parcialmente responsables, de la regulación del flujo sanguíneo local. Las sustancias vasodilatadoras, como el dióxido de carbono, el ácido láctico y los iones potasio, tienden a aumentar en los tejidos cuando el flujo sanguíneo se reduce y el metabolismo celular prosigue al mismo ritmo, o cuando el metabolismo celular aumenta súbitamente. Un aumento en la concentración de metabolitos vasodilatadores produce una vasodilatación de las arteriolas, lo que incrementa el flujo sanguíneo en los tejidos y devuelve a la normalidad los valores de concentración tisular de los metabolitos.
La teoría de la demanda de oxígeno o la teoría de la demanda de nutrientes: El oxígeno es uno de los nutrientes metabólicos necesarios para provocar la contracción muscular (con la necesidad también de otros nutrientes). Por tanto, es razonable creer que los vasos sanguíneos simplemente se relajarían en ausencia de una cantidad adecuada de oxígeno, dilatándose de forma natural. Además, el aumento de la utilización de oxígeno en los tejidos como consecuencia del aumento del metabolismo podría, en teoría, disminuir la disponibilidad de oxígeno hacia las fibras musculares lisas de los vasos sanguíneos locales, lo cual también provocaría la vasodilatación local. Los esfínteres precapilares y las metaarteriolas se abren y cierran cíclicamente varias veces por minuto, siendo proporcional la duración de las fases abiertas a las necesidades metabólicas de oxígeno en los tejidos. La apertura y el cierre cíclicos se denominan vasomotilidad.

Como el músculo liso que necesita oxígeno se mantiene contraído, se podría suponer que la fuerza de la contracción de los esfínteres aumentaría cuando aumenta la concentración de oxígeno. Por tanto, se supone que cuando la concentración de oxígeno aumenta por encima de cierto nivel los esfínteres precapilares y las metaarteriolas se cerrarán hasta que las células tisulares consuman el exceso de oxígeno. Sin embargo, una vez que se ha eliminado y que la concentración de oxígeno ha descendido lo suficiente, los esfínteres se abrirán una vez más para comenzar el ciclo de nuevo. Es decir, según los datos disponibles, la teoría de la sustancia vasodilatadora o la teoría de la demanda de oxígeno explicarían la regulación sanguínea local a corto plazo en respuesta a las necesidades metabólicas de los tejidos. Probablemente, en realidad se produzca una combinación de los dos mecanismos.
En cualquier tejido del organismo el rápido incremento de la presión arterial provoca un aumento inmediato del flujo sanguíneo. Sin embargo, en menos de 1 min ese flujo volverá a la normalidad en la mayoría de los tejidos, incluso aunque la presión arterial se mantenga elevada. Esta normalización del flujo se denomina autorregulación.
La teoría metabólica se puede entender fácilmente si se aplican los principios básicos de regulación del flujo sanguíneo local que hemos comentado en las secciones previas. Estos nutrientes (en especial, el oxígeno), junto con el descenso en los niveles tisulares de vasodilatadores, provocan entonces la constricción de los vasos sanguíneos y el retorno del flujo casi a la normalidad, a pesar de que aumente la presión.
La teoría miógena habría otro mecanismo no relacionado con el metabolismo tisular que explicaría el fenómeno de la autorregulación. Esta teoría se basa en la observación de que el estiramiento brusco de los vasos sanguíneos pequeños provoca la contracción del músculo liso de la pared vascular durante unos segundos. Por tanto, se ha propuesto que cuando una presión arterial elevada estira el vaso se provoca, a su vez, una constricción vascular reactiva que reduce el flujo sanguíneo casi a la normalidad. Por el contrario, con presiones bajas el grado de estiramiento del vaso es menor, por lo que el músculo liso se relaja, reduce la resistencia vascular y ayuda a recuperar la normalidad del flujo. La respuesta miógena es inherente al músculo liso vascular y puede producirse en ausencia de influencias nerviosas u hormonales. Es más pronunciada en las arteriolas, pero se puede ver también en arterias, vénulas, venas e incluso en vasos linfáticos. La contracción miógena se inicia por la despolarización vascular inducida por el estiramiento, que tiende a aumentar rápidamente la entrada de ion calcio desde el líquido extracelular hacia las células, provocando su contracción. Los cambios de la presión vascular también pueden abrir o cerrar otros canales iónicos que influyen en la contracción vascular. Se desconocen los mecanismos por los cuales los cambios de la presión provocan la apertura o el cierre de los canales iónicos vasculares, pero es probable que consistan en efectos mecánicos de la presión sobre las proteínas extracelulares que están ancladas en los elementos del citoesqueleto de la pared vascular o en los propios canales iónicos.
Un mecanismo clave para la regulación del flujo sanguíneo local a largo plazo consiste principalmente en cambiar la cantidad de vascularización de los tejidos. Por ejemplo, la vascularización aumenta si el metabolismo de un tejido dado aumenta durante un período prolongado, en un proceso denominado generalmente angiogenia. Es decir, se produce una reconstrucción física real de la vasculatura tisular para cubrir las necesidades de los tejidos. Esta reconstrucción es rápida (en días) en los animales muy jóvenes y también en un tejido de nuevo crecimiento, como en el tejido cicatricial o el tejido canceroso, pero es mucho más lenta en los tejidos antiguos y bien establecidos. Por tanto, el tiempo necesario para que tenga lugar la regulación a largo plazo puede ser de solo unos días en el recién nacido o hasta meses en la tercera edad. Además, el grado último de respuesta es mucho mejor en tejidos más jóvenes que en los más mayores, por lo que la vascularización se ajustará en el recién nacido, para cubrir casi exactamente las necesidades de flujo sanguíneo del tejido, mientras que en los más antiguos la vascularización va por detrás de las necesidades de los tejidos.
El oxígeno es importante no solo para el control a corto plazo del flujo sanguíneo local, sino también para el control a largo plazo. Un ejemplo es el aumento de la vascularización de los tejidos en los animales que viven en altitudes elevadas, donde el oxígeno atmosférico es bajo. En los recién nacidos prematuros que son tratados en tiendas de oxígeno con fines terapéuticos, el exceso de oxígeno provoca la interrupción casi inmediata del crecimiento vascular nuevo en la retina e incluso la degeneración de algunos de los vasos pequeños que ya se han formado.

Hay una docena o más de factores que aumentan el crecimiento de los vasos sanguíneos nuevos, siendo casi todos ellos péptidos pequeños. Los cuatro factores mejor identificados son el factor de crecimiento del endotelio vascular (VEGF), el factor de crecimiento de los fibroblastos, el factor de crecimiento derivado de plaquetas (PDGF) y la angiogenina, aislados cada uno de ellos en tejidos que tienen un aporte sanguíneo inadecuado. Presumiblemente, es la deficiencia de oxígeno tisular o de otros nutrientes la que provoca la formación de los factores de crecimiento vascular.
La angiogenia comienza con la gemación de nuevos vasos desde otros vasos. El primer paso es la disolución de la membrana basal de las células endoteliales en el punto de gemación, seguido por la reproducción rápida de las células endoteliales nuevas que buscan la salida a través de la pared del vaso en cordones que se van extendiendo directamente hacia la fuente del factor angiogénico. Las células de cada cordón continúan dividiéndose y se pliegan rápidamente formando un tubo. A continuación, este tubo se conecta con otro tubo que ha nacido de otro vaso donante (otra arteriola o vénula) y forma un asa capilar a través de la cual la sangre comienza a fluir. Si el flujo es suficientemente grande, los miocitos pequeños invaden finalmente la pared, por lo que algunos de los vasos nuevos finalmente se convertirán en arteriolas o vénulas nuevas o incluso en vasos más grandes. Es decir, la angiogenia explica la forma en que los factores metabólicos de los tejidos locales provocan el crecimiento de vasos nuevos.

Una característica especial de gran valor del control vascular a largo plazo es que la vascularización se determina principalmente por el nivel máximo de flujo sanguíneo necesario y no por la necesidad media. Por ejemplo, durante el ejercicio intenso el flujo sanguíneo de todo el cuerpo aumenta el flujo sanguíneo en reposo hasta seis u ocho veces. Este mayor exceso de flujo puede no ser necesario más que durante algunos minutos cada día, aunque esta necesidad breve provoca la formación de factores angiogénicos suficiente en los músculos para aumentar su vascularización según necesidades. Si no fuera por esta capacidad, cada vez que una persona intentara hacer un ejercicio intenso los músculos no podrían recibir los nutrientes adecuados, en especial el oxígeno necesario, por lo que los músculos no se contraerían.
En la mayoría de los tejidos, cuando se bloquea una arteria o una vena se desarrolla un canal vascular nuevo que rodea el bloqueo y permite que se vuelva a suministrar sangre al tejido afectado, al menos parcialmente. La primera etapa de este proceso es la dilatación de los bucles vasculares pequeños que ya conectan ese vaso proximal al bloqueo con el vaso distal. Esta dilatación se produce en el primer o segundo minutos, lo que indica que la dilatación está mediada probablemente por factores metabólicos. Después de esta apertura inicial de los vasos colaterales, el flujo es menor de la cuarta parte de lo necesario para cubrir todas las necesidades tisulares. No obstante, la apertura se produce en las horas siguientes, por lo que antes de 1 día pueden estar ya cubiertas la mitad de las necesidades tisulares y en pocos días el flujo sanguíneo suele ser suficiente para cubrir todas estas necesidades. Los vasos colaterales continúan creciendo durante muchos meses después, normalmente formando muchos canales colaterales pequeños en lugar de un único vaso de gran tamaño. En reposo, el flujo sanguíneo puede volver muy cerca de los valores normales, pero los nuevos canales son suficientemente grandes como para aportar el flujo sanguíneo necesario durante la actividad tisular agotadora. Es decir, el desarrollo de los vasos colaterales sigue los principios habituales del control a corto y largo plazo del flujo sanguíneo local, consistiendo el control a corto plazo en la dilatación metabólica rápida seguido crónicamente por el crecimiento e ingurgitación de los vasos nuevos en un período de semanas y meses.
En los pequeños vasos sanguíneos que se contraen como respuesta al aumento de la presión arterial, las células de músculo liso vascular y las células endoteliales se reorganizan gradualmente, en un período de unos días o varias semanas, en torno a un menor diámetro luminal, en un proceso denominado remodelación eutrófica de entrada, sin que se produzcan cambios en el área de la sección transversal de la pared vascular.
En las grandes arterias que no se contraen como respuesta al aumento de presión, la pared del vaso queda expuesta a una mayor tensión de la pared, lo que estimula una respuesta de remodelación hipertrófica y un aumento en el área en sección transversal de la pared vascular. La respuesta hipertrófica incrementa el tamaño de las células de músculo liso vascular y estimula la formación de proteínas de matriz extracelular adicionales, como colágeno y fibronectina, que refuerzan la resistencia de la pared vascular para hacer frente al aumento de las presiones arteriales. No obstante, esta respuesta hipertrófica también enrigidece los vasos sanguíneos, un signo distintivo de hipertensión crónica.
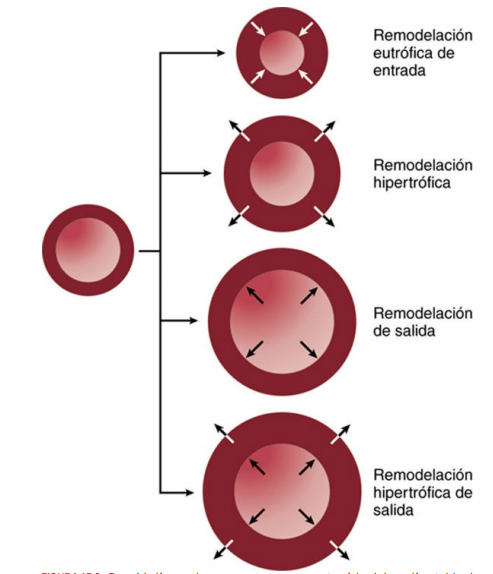
Cuando el flujo sanguíneo se reduce de forma importante, también se reduce el diámetro de la luz vascular, y cuando disminuye la presión arterial, normalmente disminuye el grosor de la pared vascular. Así pues, la remodelación vascular es una respuesta adaptativa importante de los vasos sanguíneos al crecimiento y desarrollo tisular, así como a los cambios fisiológicos y patológicos en la presión arterial y el flujo sanguíneo de los tejidos.
El control humoral de la circulación se refiere al control por las sustancias segregadas o absorbidas en los líquidos del organismo, como hormonas y factores producidos localmente. Algunas de esas sustancias se forman en glándulas especiales y se transportan en la sangre por todo el organismo, mientras que otras se forman en algunas zonas del tejido afectado y provocan solo efectos circulatorios locales. Entre los factores humorales más importantes que afectan a la función circulatoria destacan los que se describen en los siguientes apartados.
La noradrenalina es una hormona vasoconstrictora especialmente potente; la adrenalina es menos potente y en algunos tejidos provoca incluso una vasodilatación leve. Cuando se estimula el sistema nervioso simpático en el cuerpo durante el estrés o el ejercicio, las terminaciones nerviosas simpáticas de cada tejido liberan noradrenalina, que excita al corazón y contrae las venas y las arteriolas.
La angiotensina II es otra sustancia vasoconstrictora potente. Tan solo una millonésima de gramo puede aumentar la presión arterial de un ser humano en 50 mmHg o más. El efecto de angiotensina II contrae potentemente las pequeñas arteriolas. Si esta contracción sucede en un tejido aislado, el flujo sanguíneo de esa zona disminuirá mucho, aunque la importancia real de la angiotensina II es que normalmente actúa sobre muchas de las arteriolas del organismo al mismo tiempo, para aumentar la resistencia periférica total y reducir la excreción de sodio y agua en los riñones, lo que aumenta la presión arterial.
Sustancias denominadas cininas que provocan una vasodilatación potente cuando se forman en la sangre y en los líquidos tisulares de algunos órganos. Las cininas son pequeños polipéptidos que se escinden por enzimas proteolíticas a partir de α2 – globulinas del plasma o los líquidos tisulares. Una enzima proteolítica de particular importancia para tal fin es la calicreína, que se encuentra en la sangre y los líquidos tisulares en una forma inactiva. Esta calicreína inactiva se activa por la maceración de la sangre, por la inflamación tisular o por otros efectos químicos o físicos similares. A medida que se va activando la calicreína actúa inmediatamente sobre la α2 -globulina para liberar una cinina llamada calidina, que después se convierte en bradicinina gracias a las enzimas tisulares. Una vez formada, la bradicinina persiste durante solo unos minutos, porque se inactiva por la enzima carboxipeptidasa o por la enzima convertidora, la misma que participa en la activación de la angiotensina. La histamina tiene un efecto vasodilatador potente sobre las arteriolas y, como la bradicinina, puede aumentar en gran medida la porosidad capilar permitiendo la pérdida tanto de líquidos como de proteínas plasmáticas hacia los tejidos. En muchas situaciones patológicas la dilatación arteriolar intensa y el aumento de la porosidad capilar producida por la histamina provoca la pérdida de cantidades enormes de líquido desde la circulación hacia los tejidos, induciendo el edema.
