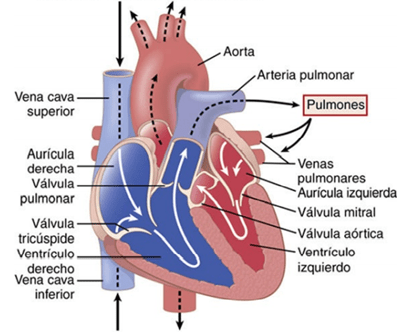
- El corazón, que se muestra está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente. Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre: 1) hacia la circulación pulmonar por el ventrículo derecho, o 2) hacia la circulación sistémica por el ventrículo izquierdo.
- El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor. Las fibras especializadas de excitación y de conducción del corazón se contraen solo débilmente porque contienen pocas fibrillas contráctiles, en cambio, presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco.
Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos intercalados, realmente son membranas celulares que separan las células musculares cardíacas individuales entre sí. Es decir, las fibras musculares cardíacas están formadas por muchas células individuales conectadas entre sí en serie y en paralelo. En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión. Por tanto, desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas. El corazón realmente está formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
El potencial de acción registrada en una fibra muscular ventricular es de 105 mV en promedio, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción ha- ce que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
- Fase 0 (despolarización), los canales de sodio rápidos se abren. Cuando la célula cardíaca es estimulada y se despolariza, el potencial de membrana se hace más positivo. Los canales de sodio activados por el voltaje (canales de sodio rápidos) se abren y permiten que el sodio circule rápidamente hacia el interior de la célula y la despolarice. El potencial de membrana alcanza +20 mV aproximadamente antes de que los canales de sodio se cierren.
- Fase 1 (repolarización inicial), los canales de sodio rápidos se cierran. Los canales de sodio se cierran, la célula empieza a repolarizarse y los iones potasio salen de la célula a través de los canales de potasio. Fase 2 (meseta), los canales de calcio se abren y los canales de potasio rápidos se cierran. Tiene lugar una breve repolarización inicial y el potencial de acción alcanza una meseta como consecuencia de: 1) una mayor permeabilidad a los iones calcio, y 2) la disminución de la permeabilidad a los iones potasio. Los canales de calcio activados por el voltaje se abren lentamente durante las fases 1 y 0, y el calcio entra en la célula. Después, los canales de potasio se cierran, y la combinación de una reducción en la salida de iones potasio y un aumento de la entrada de iones calcio lleva a que el potencial de acción alcance una meseta.
- Fase 3 (repolarización rápida), los canales de calcio se cierran y los canales de potasio lentos se abren. El cierre de los canales iónicos de calcio y el aumento de la permeabilidad a los iones potasio, que permiten que los iones potasio salgan rápidamente de la célula, pone fin a la meseta y devuelve el potencial de membrana de la célula a su nivel de reposo.
- Fase 4 (potencial de membrana de reposo) con valor medio aproximado de −90 mV.
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s en la mayoría de las partes del sistema, lo que permite una conducción razonablemente rápida de la señal excitadora hacia las diferentes partes del corazón.
El músculo cardíaco, al igual que todos los tejidos excitables, es refractario a la reestimulación durante el potencial de acción. El período refractario normal del ventrículo es de 0,25 a 0,30 s, que es aproximadamente la duración del potencial de acción en meseta prolongado. Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo normal excitar el músculo, pero, sin embargo, se puede excitar con una señal excitadora muy intensa. El período refractario del músculo auricular es mucho más corto que el de los ventrículos (aproximadamente 0,15 s para las aurículas, en comparación con 0,25 a 0,30 s para los ventrículos).
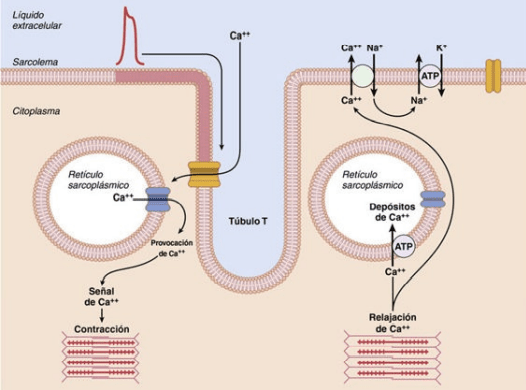
- El término acoplamiento excitación-contracción se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan. Cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T). Los potenciales de acción de los túbulos T, a su vez, actúan sobre las membranas de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia el sarcoplasma muscular desde el retículo sarcoplásmico. En algunas milésimas de segundo más estos iones calcio difunden hacia las miofibrillas y catalizan las reacciones químicas que favorecen el deslizamiento de los filamentos de actina y de miosina entre sí, lo que da lugar a la contracción muscular. El calcio que entra en la célula activa después los canales de liberación de calcio, también denominados canales de receptor de rianodina, en la membrana del retículo sarcoplásmico, para activar la liberación de calcio en el sarcoplasma. Los iones calcio en el sarcoplasma interaccionan después con la troponina para iniciar la formación y contracción de puente transversal. La fuerza de la contracción del músculo cardíaco depende en gran medida de la concentración de iones calcio en los líquidos extracelulares. La duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.
Ciclo cardíaco
Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal. Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o 0,833 s por latido. Cuando aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye, incluidas las fases de contracción y relajación. La duración del potencial de acción y el período de contracción (sístole) también decrece, aunque no en un porcentaje tan elevado como en la fase de relajación (diástole). Para una frecuencia cardíaca normal de 72 latidos/min, la sístole comprende aproximadamente 0,4 del ciclo cardíaco completo. Para una frecuencia cardíaca triple de lo normal, la sístole supone aproximadamente 0,65 del ciclo cardíaco completo. Esto significa que el corazón que late a una frecuencia muy rápida no permanece relajado el tiempo suficiente para permitir un llenado completo de las cámaras cardíacas antes de la siguiente contracción. La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. Por tanto, las aurículas actúan como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%. Sin embargo, el corazón puede seguir funcionando en la mayor parte de las condiciones incluso sin esta eficacia de un 20% adicional porque normalmente tiene la capacidad de bombear entre el 300 y el 400% más de sangre de la que necesita el cuerpo en reposo.
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
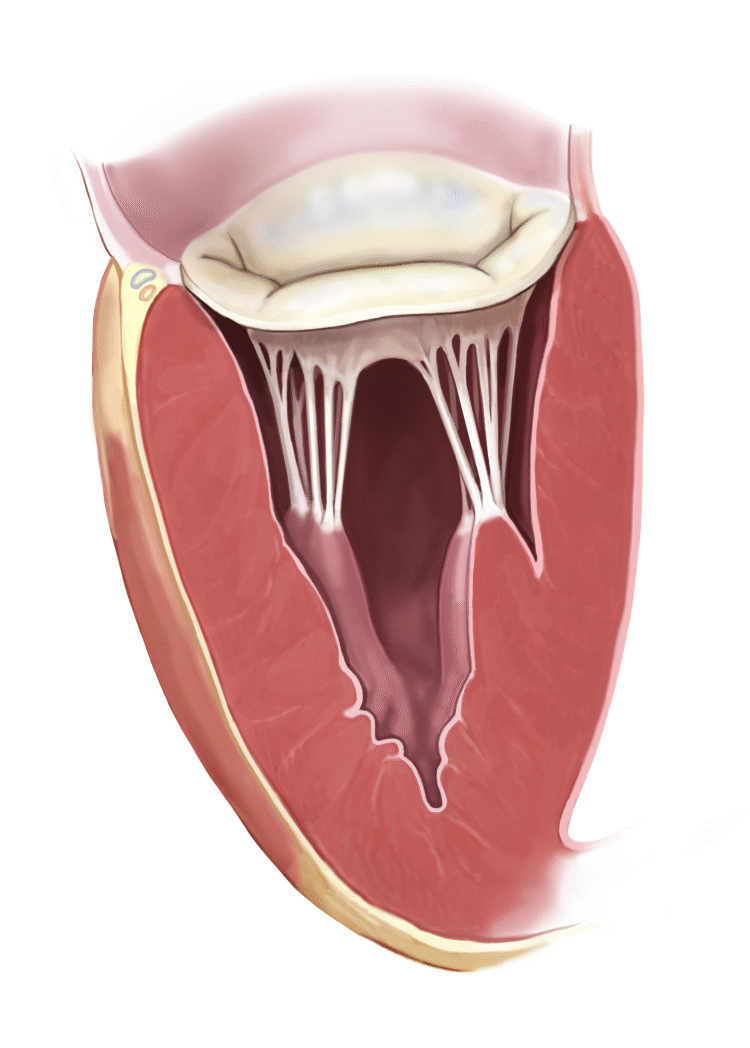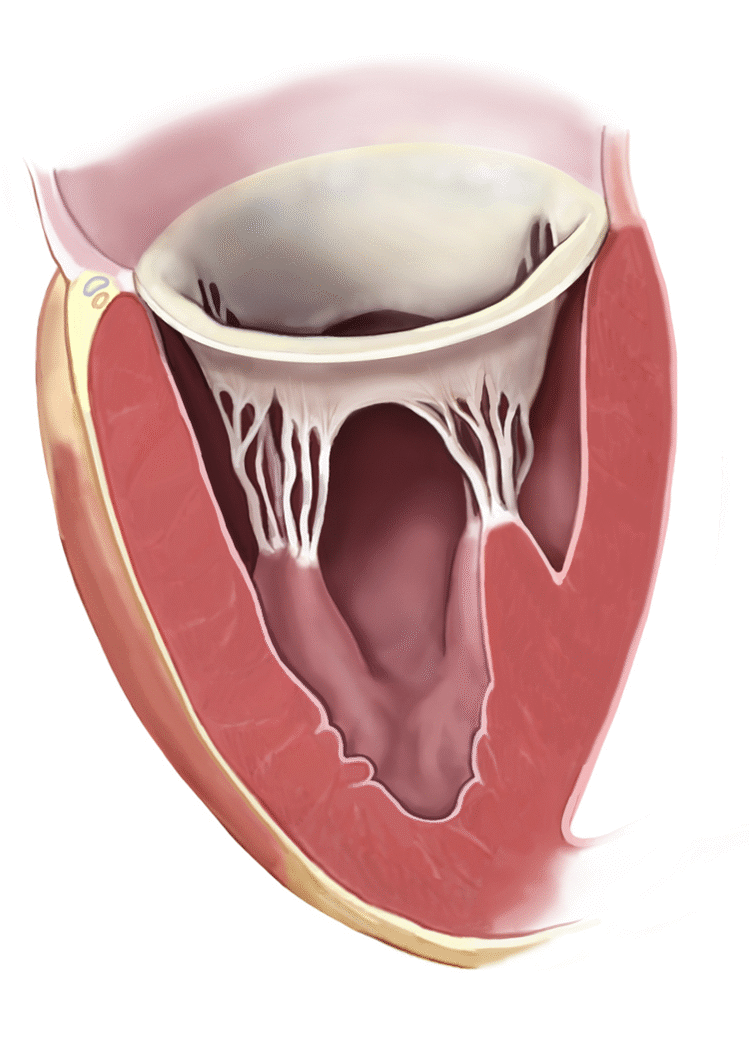
Los músculos papilares se contraen cuando se contraen las paredes ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
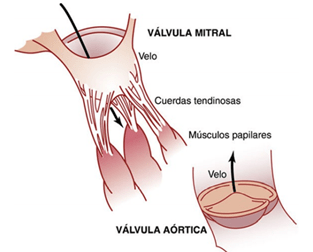
Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares.

Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. La entrada de sangre en las arterias durante la sístole hace que sus paredes se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Al final de la sístole, después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una presión elevada en las arterias, incluso durante la diástole. Se produce una incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado. Después de que se haya cerrado la válvula aórtica, la presión en el interior de la aorta disminuye lentamente durante toda la sístole porque la sangre que está almacenada en las arterias elásticas distendidas fluye continuamente a través de los vasos periféricos de nuevo hacia las venas. Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente ha disminuido hasta aproximadamente 80 mmHg (presión diastólica), que es dos tercios de la presión máxima de 120 mmHg (presión sistólica) que se produce en la aorta durante la contracción ventricular.
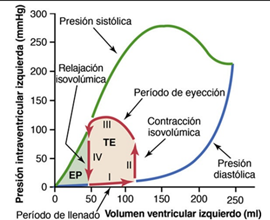

La curva de presión diastólica se determina llenando el corazón con volúmenes de sangre progresivamente mayores y midiendo la presión diastólica inmediatamente antes de que se produzca la contracción ventricular, que es la presión telediastólica del ventrículo. La curva de presión sistólica se determina registrando la presión sistólica que se alcanza durante la contracción ventricular a cada volumen de llenado. Hasta que el volumen del ventrículo que no se está contrayendo no aumenta por encima de aproximadamente 150 ml, la presión «diastólica» no aumenta mucho. Por tanto, hasta este volumen la sangre puede fluir con facilidad hacia el ventrículo desde la aurícula. Por encima de 150 ml la presión diastólica ventricular aumenta rápidamente, en parte porque el tejido fibroso del corazón ya no se puede distender más y en parte porque el pericardio que rodea el corazón se ha llenado casi hasta su límite. Durante la contracción ventricular, la presión «sistólica» aumenta incluso a volúmenes ventriculares bajos y alcanza un máximo a un volumen ventricular de 150 a 170 ml.
Control del corazón por los nervios simpáticos y parasimpáticos
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón.
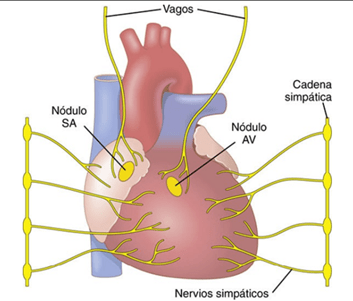
La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min. Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces, además del aumento del gasto que produce el mecanismo de Frank-Starling que ya se ha comentado. Por el contrario, la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo aproximadamente un 30% por encima del que habría sin estimulación simpática. Por tanto, cuando la actividad del sistema nervioso simpático disminuye por debajo de lo normal, tanto de la frecuencia cardíaca como la fuerza de la contracción del músculo ventricular se reducen, con lo que disminuye el nivel de bombeo cardíaco hasta un 30% por debajo de lo normal. La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%. Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón. Esta distribución explica por qué el efecto de la estimulación vagal tiene lugar principalmente sobre la reducción de la frecuencia cardíaca, en lugar de reducir mucho la fuerza de la contracción del corazón. Sin embargo, la gran disminución de la frecuencia cardíaca, combinada con una ligera reducción de la fuerza de la contracción cardíaca, puede reducir el bombeo ventricular en un 50% o más.
